Morfogene gradienter og form-dannelse
Av Otangelo Grasso, oversatt herfra
Å etablere et dyrs kroppsplan avhenger av mange mekanismer som går utover genetisk informasjon.
Utviklingen av en organisme fra en udifferensiert enkeltcelle, til en romlig kompleks struktur krever romlig mønstring av celle skjebner , på tvers av vev. Flere forskjellige signalveier er avgjørende for dannelse av organisert vev, organisme form og arkitektur. Romlig mønster av et utviklende dyr krever at celler blir forskjellige i henhold til deres posisjoner i embryoet, noe som betyr at celler må svare på ekstracellulære signaler produsert av andre celler, spesielt deres naboer. I det som sannsynligvis er den vanligste modusen for romlig mønstring, starter en gruppe pluripotente celler med det samme utviklingspotensialet, og et signal fra celler utenfor gruppen induserer deretter ett eller flere medlemmer av gruppen, for å endre sin karakter. Noen induktive signaler avhenger av celle -celle -kontakt; Andre opptrer over et lengre område og er formidlet av molekyler som diffunderer gjennom det ekstracellulære mediet eller blir transportert i blodomløpet. De fleste av de kjente hendelsene i dyreutvikling styres av et lite antall signalveier.
Et kjernesett med viktige morfogener regulerer utvikling. Morfogener signaliserer molekyler som danner konsentrasjonsgradienter med lang rekkevidde, som mønstrer et felt av celler. Etter at morfogener er utskilt fra en gruppe celler, blir de transportert over vev for å etablere en gradient. Transporten av morfogener er mediert av kinesin- og dyneinmotorproteiner langs mikrotubulens nettverk av aksoner. Morfogener i nærheten av den lokale kilden blir festet til skyttelen og morfogen-skyttel-komplekset transporteres over vev. Deretter blir morfogen-skyttel-komplekset degradert, noe som resulterer i at morfogenet blir lokalt immobilisert, og begynner å danne en lokal gradient. Bioelektriske signaler og hvilemembran-potensialer over vev, er også, og i tillegg, avgjørende for riktig mønster i flere organismer. Videre er det også nevrotransmittere (kjemiske budbringere som overfører en melding fra en nervecelle over synapsen til en målcelle) etablerer venstre-høyre mønster i embryoer gjennom regulering av ionestrømmer. Kalsium (Ca2+) signalering spiller også en nøkkelrolle i å formidle cellulære responser på morfogener. I tillegg til å formidle cellulær differensiering og spredning, påvirker Ca2+ -signaler også cellulær migrasjon, som respons på morfogengradienter.
Robusthet er et allestedsnærværende trekk ved biologiske systemer som sikrer at spesifikke funksjoner i systemet opprettholdes til tross for eksterne og interne forstyrrelser. Videre er retningsbestemt transport av stoffer gjennom nervesystemet nødvendig for å oppnå skalafri morfogenmønstring og bestemmelse av kroppsaksen.
 Celler føler og tolker deres posisjon som en funksjon av mengden signal de mottar, og oppnår dermed 'posisjonsinformasjon'. Innenfor større vev gir nevrale nettverk rettet informasjon, via fysiologisk signalering, som supplerer posisjonsinformasjon gjennom diffusjon. Denne posisjons-informasjonen spesifiserer genuttrykk og påfølgende skjebner av celler i vev.
Celler føler og tolker deres posisjon som en funksjon av mengden signal de mottar, og oppnår dermed 'posisjonsinformasjon'. Innenfor større vev gir nevrale nettverk rettet informasjon, via fysiologisk signalering, som supplerer posisjonsinformasjon gjennom diffusjon. Denne posisjons-informasjonen spesifiserer genuttrykk og påfølgende skjebner av celler i vev.
Noen få nøkkelsentenser: Celler opererer etter biologiske designregler - Utviklingsceller har et ordforråd - det er intercellulær kommunikasjon - celler kommuniserer med hverandre gjennom celle -celle gap -kryss, som treningsmodulasjon, som er kontroll, styrer, induserer og regulerer mønster dannelse. - Det er såkalt fysiologisk signalering, og det er positive diffusjons-regulatorer - det er også signaler som formerer modulering av ionflukser gjennom spenningsrettede ionekanaler - det er signalmolekyler som gir informasjon for retningssignaler - det er også bioelektriske signaler - Celler vet hvordan de skal tolke sin posisjon ved å skaffe 'posisjonsinformasjon' gjennom et koordinat-system, som er modulert og mediert gjennom morfogener som tillater dannelse av spesifikke mønstre gjennom gradienter. Disse regulerer utviklingen; De spesifiserer også celleskjebner.
Det er en generell betydning av mekaniske krefter i regulering av vekst og morfogenese. Mekaniske krefter, cellulær mekanikk og vevsmekanikk er viktige komponenter i morfogenese og kan påvirke mønsterdannelse.
Som et generelt opplegg kan det observeres at det ikke bare er kjemiske reaksjoner, men molekyler og celler opererer, oppfører seg, migrerer, plasserer seg, organiserer, danner vev, spesialiserer seg basert på instruksjoner gjennom forskjellige signalnettverk og mekanismer, som har som kilde forskjellige epigenetiske signalveier. Integrerte, nettverksbaserte informasjonssystemer tillater lite, om noen, feil, noe som utvilsomt fører til sykdom. Dette forhindres av robusthet som må settes opp fra begynnelsen. Dette fører til min (Grassos) forståelse av intelligent design som den beste forklaringen.
BILDE 1: Morfogengradienter og mønsterdannelse (1)
Lite antall konserverte celle -celle signalveier koordinerer romlig mønster-dannelse
Romlig mønster av et utviklende dyr, krever at celler blir forskjellige i henhold til deres posisjoner i embryoet, noe som betyr at celler må svare på ekstracellulære signaler produsert av andre celler, spesielt deres naboer. I det som sannsynligvis er den vanligste modusen for romlig mønstring, starter en gruppe celler med samme utviklingspotensiale, og et signal fra celler utenfor gruppen får deretter ett eller flere medlemmer av gruppen til å endre sin karakter. Denne prosessen kalles induktiv signalering. Generelt er det induktive signalet begrenset i tid og rom, slik at bare en delmengde av cellene som er i stand til å svare - cellene nær kilden til signalet - reagerer på det induserte tegnet
BILDE 2: Induktiv signalering celleavstand

Induktiv signalering.
Noen induktive signaler avhenger av celle -celle -kontakt; Andre opptrer over et lengre område og er formidlet av molekyler som diffunderer gjennom det ekstracellulære mediet eller blir transportert i blodomløpet. De fleste av de kjente induktive hendelsene i dyreutvikling styres av et lite antall høyt bevarte signalveier, inkludert:
transformerende vekstfaktor-beta (TGFβ), Wnt, Hedgehog, V-formede kutt, reseptor tyrosinkinase (RTK) -veier
Oppdagelsen av det begrensede ordforrådet som utviklende celler bruker for intercellulær kommunikasjon, har dukket opp de siste 25 årene som et av de store forenklende trekk ved utviklingsbiologi.
Romgrenser er kildene til morfogener. Morfogener signaliserer molekyler som produseres fra en lokalisert kilde som danner konsentrasjonsgradienter med lang rekkevidde som gir mønster til et felt med celler (figur 7A). Celler tolker deres posisjon som en funksjon av mengden signal de mottar, og oppnår dermed 'posisjonsinformasjon' (Wolpert, 1989, 1996). Signalmolekyler må oppfylle to strenge kriterier for å kvalifisere som morfogener: (1) Effekten deres må utøves på en konsentrasjonsavhengig måte og (2) de må virke direkte på målceller i en avstand fra kilden (dvs. ikke gjennom en sekundær relémekanisme). Når disse kriteriene er oppfylt, kan den lokale konsentrasjonen tolkes som et mål på avstand fra kilden til signalet (figur 7A). De tre signalproteiner som kvalifiserer som morfogener i Drosophila vingen -utvikling er Hedgehog (HH), Decapentaplegic (DPP) og Wingless (WG) (f.eks. WG er vist i figur 7).
Figur 7 Eksempel på en morfogengradient: Bilde 3. Wingless (WG).

(a) Tegning av et avsnitt av vingepose-epitel (øvre panel) og av vingeposen i en apikal utsikt (nedre panel). Cellene i sentrum uttrykker WG som genererer en gradient over det tilstøtende vevet. Celler på forskjellige avstander får forskjellige nivåer av WG.
(B) Skjematisk tegning av WG -gradienten og målgenaktiveringen i epitelet som respons på WG (øvre panel). WG -gradienten aktiverer målgenene etterpåklokskap (Cyan) og Distalless (blå) på forskjellige terskelnivåer. Ekspresjonsdomenene til Hindsight og Distalless er avbildet i de samme fargene i det nedre panelet. (c) WG nedregulerer sin egen reseptor Drosophila Frizzled 2 (DFZ2; Blue), former dens gradient og gjengir celler i en avstand fra kilden til WG mer følsomme.
Det er generelt akseptert at celler tolker gradienten ved å fremkalle differensielle transkripsjonsresponser, avhengig av konsentrasjonen av morfogener de blir utsatt for. Dette krever at celler tar beslutninger avhengig av forskjellige terskelnivåer av signalveisaktivitet. Konseptet innebærer at en enkelt hendelse - nemlig produksjon av et utskilt molekyl ved en lokalisert kilde - kan føre til dannelse av flere forskjellige celletyper i et riktig romlig forhold til hverandre (figur 7). Dette representerer en svært effektiv måte å generere komplekse mønstre i tidligere ikke-involverte celler (Gjennomgang fra: Gurdon og Bourillot, 2001).
Ulike moduser for morfogenbevegelse/transport er blitt påberopt for å forklare dannelse av langdistanse: ekstracellulær diffusjon av det utskilte molekylet, sykluser av reseptormediert endocytose og resecresjon (plan transcytose), membraner (argosomer) og cytoplasmisk utvidelse (argosomer) . Selv om det hittil ikke er overbevisende bevis for at andre mekanismer enn diffusjon bidrar produktivt til gradientdannelse, er det absolutt sannsynlig at disse mekanismene kan gjøre det. Cytonemes og argosomer eksisterer, men har ennå ikke vist seg å være påkrevd for å formidle ligandbevegelse eller for å overføre signaler. Bevis presentert til fordel for endocytose og resettering som en mekanisme for ligandtransport har blitt stilt spørsmål ved. Ytterligere eksperimentelle bevis er nødvendig for å støtte eller tilbakevise disse mulighetene. Flere faktorer har blitt involvert i å forme morfogengradienter: Posttranslasjonell modifisering av liganden kan påvirke diffusibilitet (f.eks. Acylering av HH og WG (Ingham, 2000, 2001)). Regulering av reseptornivåer kan påvirke ligandbevegelsen (figur 7C). Celleoverflate heparan-sulfatproteoglykaner og utskilt enzymer som modifiserer dem, kan modulere ligand bevegelse. Utskilt ligandbindende proteiner kan påvirke bevegelse eller stabilitet. I tillegg kan cellulære responser på disse ligandene også moduleres, og bidra til å forme aktivitetsgradientene, i stedet for ligandgradientene per se.
Samspill mellom morfogen -styrt posisjonsinformasjons-systemer og fysiologisk signalering 3. desember 2019 (4)
Utviklingen av en organisme fra en udifferensiert enkeltcelle til en romlig kompleks struktur krever romlig mønstring av celle-skjebner over vev. Posisjonsinformasjon, foreslått av Lewis Wolpert i 1969, har ført til karakterisering av mange komponenter, involvert i regulering av morfogensignaleringsaktivitet. Kvantitative og systembaserte tilnærminger er i økende grad nødvendig for å definere generelle biologiske designregler, som styrer posisjonsinformasjonssystemer for å utvikle organismer. Det er forskjellige roller for fysiologisk signalering i å modulere og formidle morfogenbasert mønsterdannelse. Likheter mellom nevral overføring og morfogenbaserte mønsterdannelsesmekanismer, antyder underliggende delte prinsipper for aktiv cellebasert kommunikasjon. Innenfor større vev gir nevrale nettverk rettet informasjon, via fysiologisk signalering, som supplerer posisjonsinformasjon gjennom diffusjon. Videre viser montering av bevis at fysiologisk signalering spiller en rolle i å sikre robusthet av morfogenbasert signalering. Vi avslutter med å fremheve flere fremragende spørsmål angående fysiologisk signalering i morfogenbasert mønsterdannelse. å belyse hvordan fysiologisk signalering påvirker posisjonsinformasjon er kritisk for å forstå den nære koblingen av utviklings- og cellulære prosesser i sammenheng med utvikling, sykdom og regenerering. (2)
Lewis Wolpert foreslo posisjonsinformasjon som en mekanisme der differensialgenuttrykk av celler resulterer i romlige mønstre av cellulær differensiering. Innenfor begrepet posisjonsinformasjon definerer et koordinatsystem størrelsen og retningen til den posisjonsinformasjonen som føles av celler. Denne posisjonsinformasjonen spesifiserer genuttrykk og påfølgende skjebner av celler i vev (figur nedenfor)
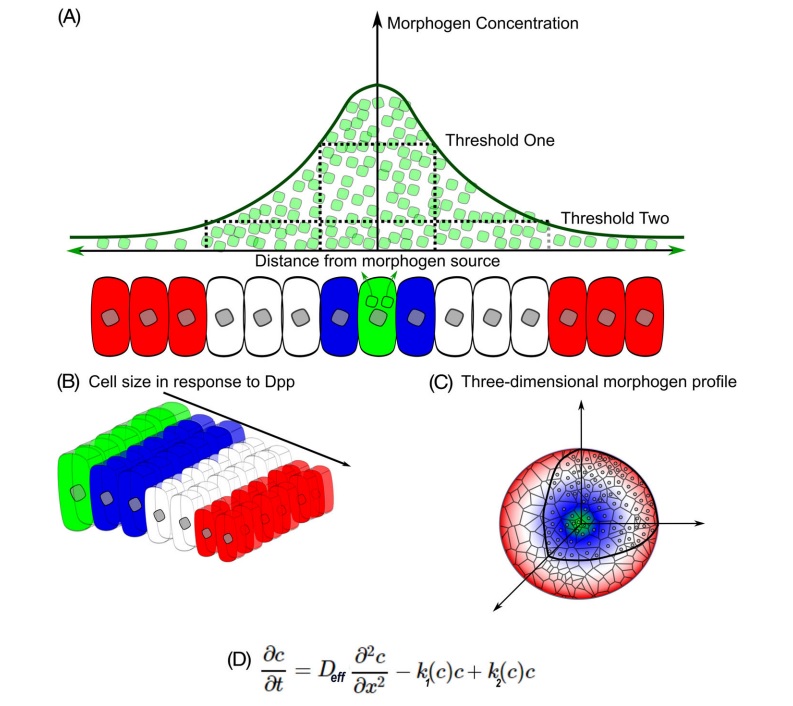
A, klassisk syn på posisjonsinformasjon og morfogenese.
Det klassiske synet på posisjonsinformasjon stammer fra den franske flaggmodellen, der kildeceller utskiller morfogener som transporteres gjennom et vev på grunn av diffusjon. A, morfogenmolekyler (grønt) skilles ut fra kildeceller der morfogenkonsentrasjon vil være den høyeste. Morfogener reiser til nabokeller for å etablere en konsentrasjonsgradient. Celler vil ha differensielle biologiske responser på morfogengradienten avhengig av flere terskelnivåer. Blå celler er lokalisert der morfogenkonsentrasjoner er over den høyere terskelen. Hvite celler føler morfogenkonsentrasjoner som er over den nedre terskelen. Røde celler oppdager lave til ingen morfogener.
B, celler reagerer på morfogener for å bestemme cellens skjebner og cellemorfologi i en doseavhengig respons.
C, progenitorcellers evne til å skape morfogen-gradientbaserte mønstre er avhengig av vevsgeometri, størrelse, tidsmessig signalering av morfogener.
D, styrende diffusjonsligning av morfogenkonsentrasjonen i samsvar med Ficks andre diffusjonslov med en ikke -lineær nedbrytningsprofil (K1), et kildekontroll avhengig av beliggenhet (K2) og effektiv diffusjon av molekylet (DEFF). Dette resulterer i et PowerLaw-forhold der gradienten er posisjonsavhengig og gjenspeiler den lokale brattheten i gradienten.
BILDE 4. Morgogen konsentrajon
Etter Wolpert foreslo Gierer og Meinhardt modeller av morfogenfordeling for å demonstrere at relativt enkle molekylære mekanismer kan forklare dannelsen av et romlig mønstret vev. Etterfølgende eksperimentelt arbeid har vist at et kjernesett med morfogener genererer og videresender posisjonsinformasjon til celler for direkte å indusere cellulære responser basert på cellenes plassering. Fordi morfogener spiller avgjørende roller under spesifikasjonen av celle-skjebner, bidrar de til mange aspekter ved utvikling. Et kjernesett med viktige morfogener regulerer utvikling. Eksempler inkluderer medlemmer av Hedgehog (HH) -familien som er involvert i Drosophila appendage -formasjon og utvikling av kyllinger. Som et annet eksempel bidrar Wingless (WG)/INT-1 (WNT) proteiner til Drosophila appendage-utvikling og menneskelige degenerative sykdommer. BenMorfogenetiske Proteiner (BMPs), slik som Drosophila Decapentaplegic (DPP), brukes i dorsal-ventral mønstring av Drosophila-embryo og mønsterdannelse og vekstkontroll av lem primordia. BMPs regulerer også dannelsen av tidlige kimlag av pattedyr. Morfogensignalering inkluderer formidling av posisjonsinformasjon, og deres evne til å indusere mønsterdannelse gjennom gradienter. Ett morfogen kan kontrollere uttrykket av et annet under morfogenetiske prosesser, for eksempel HH-indusert DPP-aktivitet i utviklingen av Drosophila og HH som deltar i smh. med Wnt i kreft. På den annen side har beregningsmodellering vist seg å være kritisk for å forklare stadig mer komplekse datasett og motintuitive resultater fra genetiske forstyrrelser til morfogenetiske mønstringssystemer. Ytterligere beregningsinnsats har avdekket nervesystemets rolle i å tilrettelegge for regenerering i planaria. Dette, blant mange andre studier, støtter en nær analogi mellom embryonal mønstring og hjernelignende signalbehandling. Imidlertid forblir den underliggende kinetikken og dynamikken i gradientdannelse og vedlikehold i mange tilfeller dårlig forstått.
Morfogenbasert signalering under utvikling krever aktive cellulære og fysiologiske prosesser, som det er blitt bemerket gjennom analyse av viktigheten av lipidmetabolisme i morfogentransport. Her betegner vi begrepet fysiologisk signalering for å representere celleleguleringsmekanismer på ionisk og molekylært nivå som kontrollerer nøkkelcellefunksjoner som menneskehandel gjennom ekso- og endocytose, metabolske prosesser for cellulær vekst og inndeling, eller regulering av cellemekanikk. I økende grad demonstrerer bevis den sentrale rollen til fysiologiske signalhendelser i formidling av morfogenaktivitet og nye paralleller mellom nevrotransmisjon og morfogentransport over ikke-nevrale vev. Spesielt fremhever vi de funksjonelle rollene til sekundære budbringere, for eksempel kalsium (Ca2+), i formidling av morfogensekresjon, transport, nedstrøms informasjonsbehandling og gir robusthet for posisjonsinformasjon.
Cellulær mekanikk påvirker direkte mønsterdannelse og morfogengradienter
Tidlige studier av morfogenese antydet forestillingen om at kjemisk-baserte morfogengradienter er de primære signalene for mønstercelledifferensiering. Imidlertid er det nye bevis som viser at mekaniske hendelser også kan være en primær trigger i mønsterdannelse. Mekaniske krefter driver cellulær selvorganisering og strukturelle omorganiseringer av en fjærfollikkels form og genuttrykk i kyllingembryoer. Spesielt initierte mekanisk aktivering av beta-catenin nedstrøms follikkelgenekspresjon av BMP2, en nøkkelkomponent i TGF-beta-signalveien. Denne studien fungerer dermed som et slående eksempel på den generelle betydningen av mekaniske krefter i å regulere vekst og morfogenese. Mekaniske krefter, cellulær mekanikk og vevsmekanikk er viktige komponenter i morfogenese og kan påvirke mønsterdannelse. De underliggende mekanismene der mekanikk gir posisjonsinformasjon til celler, krever ytterligere belysning.
Morfogenkilder er dynamisk og kontrollert av fysiologisk signalering
Den konseptuelle franske flaggmodellen, en ikonisk beskrivelse av posisjonsinformasjon foreslått av Wolpert, spesifiserer at morfogener skilles ut fra en klynge av celler og danner en gradert distribusjon gjennom vevet for å spesifisere flere celletyper avhengig av konsentrasjonen som er senset av celler (figur A ovenfor ). Innenfor denne modellen er sekresjonen av morfogener fra kildeceller, det første trinnet i dannelsen av posisjonsinformasjon (figur B over). Dette innledende rammeverket har siden utvidet seg fra et statisk synspunkt til å omfatte dynamiske endringer i størrelsen, geometrien, beliggenheten, mekanikken og tidsmessig signalering av morfogensekreterende celler (figur C ovenfor). Et eksempel på en dynamisk morfogenkilde oppstår under morfogenesen av rygg- vedheng av Drosophila melanogaster eggeskall. De morfologiske grensene for disse strukturene, avhenger av den romlige mønstrings transkripsjonsfaktoren bredt R, som er regulert av den epidermale vekstfaktorreseptoren (EGFR) signalvei gjennom et tilbakemeldings-regulerende nettverk. Mønsteret av Broad er etablert når Gurken, en EGFR-ligand, skilles ut fra den underliggende oocytten som danner en bakre-til-fremre gradient. Senere dannes en dorsoventral gradient etter translokasjon av oocyttkjernen til den dorsale fremre cortex.41-43 Dette antyder at morfogenkilder er spatiotemporalt dynamisk og ikke alltid holder seg til rammen for passiv diffusjonsbasert transport fra en stasjonær kilde (figur D ). Denne beregningsbaserte analysen av hvordan påfølgende runder med EGFR-aktivering avgrenser romlige mønstre ble eksperimentelt bekreftet. Gitt potensialet for spatiotemporalt dynamiske morfogenkilder, kan sekresjonsmekanismer for morfogener gi innsikt i hvordan dette er mulig.
Kildeceller utskiller morfogener gjennom eksocytose, en aktiv transportmekanisme av molekyler til cellemembranen gjennom vesikler. Fusjon av vesikler til cellemembranen frigjør morfogenet. Derfor påvirkes morfogensekresjon av fysiologisk regulering av eksocytose. For eksempel regulerer kanaler innvendig kalium (IRK) kanaler frigjøring av DPP og cytosolisk Ca2+ i den utviklende Drosophila -vingen Imaginal Disc45 (figur nedenfor).
BILDE 5 Dpp-uttrykk

IRK -kanaler regulerer DPP -sekresjon og cytosolisk Ca2+.
I Drosophila -vingeskiven regulerer indre utbedring av kaliumkanaler (IRK) både sekresjonen av benmorfogen protein DPP og cytosolisk Ca2+ -nivå. Ekspresjon av DPP langs dorsal-ventral (D-V) aksen i vingeskiven er indikert i rødt. DPP-uttrykk er ikke til stede langs den fremre-posterior (A-P) aksen til vingeskiven. Inhibering av IRK-kanaler ved bruk av dominerende negative IRK2-mutanter (IRK2DN) reduserte varigheten og amplituden til Ca2+ -svingninger i DPP-produserende celler (gul kvadrat indikerer region av interesse). Videre økte tap av IRK2-kanalfunksjon baseline-konsentrasjonsnivået til DPP-GFP, mens de samtidig reduserte diskrete DPP-frigjøringshendelser. Dermed endrer IRK2 -hemming i vingeskiver samtidig intracellulær Ca2+ -aktivitet og DPP -frigjøring
IRK -kanaler letter strømning av kalium (K+) inn i cellen og har en grunnleggende rolle i å gjenopprette membranpotensialet til hvilepotensialet. Inhibering av IRK -kanaler i DPP -produserende celler, redusert DPP -sekresjon uavhengig av DPP -ekspresjon. IRK -kanaler regulerer vesikkelfrigjøring ved å endre membran-potensial mens de også endrer intracellulær Ca2+ -dynamikk. En potensiell forklaring på lignende utfall i DPP og Ca2+ etter IRK -kanal hemning, er at intracellulær Ca2+ -dynamikk regulerer DPP -frigjøring, selv om hvorvidt Ca2+ har en årsakende effekt krever ytterligere studier. Ca2+ er avgjørende for regulering av epitelfysiologi under utvikling. Hvorvidt Ca2+ direkte regulerer DPP -sekresjon er foreløpig ukjent. Gitt at feilregulering av Ca2+ ofte er assosiert med genetiske lidelser i utviklingsmessige lidelser som resulterer i neoplasmer, er det nødvendig å ytterligere karakterisere rollene til Ca2+ og andre budbringere i morfogentransport og morfogenese.
Paralleller mellom nevrotransmisjon og morfogenbaserte transportsystemer
Etter at morfogener er utskilt fra en gruppe celler, blir de transportert over vev for å etablere en gradient. Hvordan morfogener sprer og danner gradienter, er fremdeles under debatt, til tross for fremgang med å forstå de molekylære mekanismene for morfogentransport gjennom avbildningsstudier og biofysiske målinger. Flere morfogen-transportmodeller er blitt foreslått i litteraturen. Disse spenner fra passive mekanismer, for eksempel fri eller hindret diffusjon (første figur D), til cellebasert spredning ved transcytose eller cytonemer. Flere transportmekanismer kan være involvert, og dette varierer sannsynligvis på tvers av utviklings-sammenhenger. Den enkleste mekanismen for morfogentransport er passiv diffusjon der molekyler sprer seg ved tilfeldig bevegelse. Imidlertid fanger denne modellen ikke fullt ut kompleksiteten i morfogen-transportdynamikk på grunn av bevis som viser at en enkelt kilde til morfogen, ikke alltid er tilstrekkelig til å etablere en gradient. For eksempel, under Drosophila Wing Disc -utvikling, uttrykkes DWNT6 i et identisk mønster med WG, 57 mens begge BMP -ligander GBB og DPP er nødvendige for å etablere riktige morfogengradienter. Ved tilrettelagt diffusjon er morfogener stort sett immobile til de binder seg til en positiv diffusjonsregulator som forbedrer bevegeligheten. Skyttel-virksomhet er et spesielt tilfelle av tilrettelagt diffusjon, der molekylær skyttelbuss, ikke morfogener, genereres fra en lokalisert kilde. Morfogener i nærheten av den lokale kilden blir festet til skyttelen og morfogen-skyttel-komplekset transporteres over vev. Deretter blir morfogen-skyttel-komplekset degradert, noe som resulterer i at morfogenet blir immobilisert til stasjonære negative diffusjons-regulatorer og dannelsen av en gradient.
Utover ekstracellulær-diffusjonsbasert morfogentransport, kan morfogener også transporteres gjennom cellebaserte mekanismer via transcytose eller langs cellulære utvidelser kjent som Cytonemes. I tilfelle av transcytose blir signalmolekyler tatt opp av celler gjennom endocytose og frigjøres deretter gjennom eksocytose i det ekstracellulære rommet. Endocytose er prosessen der lastemolekyler, inkludert morfogener, blir absorbert og distribuert til en serie endosomer med distinkte fysiske og biokjemiske egenskaper. Et endosoms dynamikk, som fisjon, fusjon og modning, påvirker dens evne til å sortere og konsentrere lastemolekyler. For tilfeller av morfogener er endosomdynamikk dermed i stand til å etablere konsentrasjonsgradienter og signaliseringsaktiviteter på tvers av vev. Det gjenstår at det gjenstår det som er den dominerende modusen for morfogentransport i en gitt utviklingskontekst. Når det gjelder Drosophila -vingeskiven, kan DPP bevege seg gjennom regioner som er mutant for type I som transformerer vekstfaktor beta reseptor TyKkeVener (TKV) og nedstrøms transkripsjonell repressor (brinker). Dette er betydelig, gitt at støtte for transcytose stammer fra eksperimenter ved bruk av DPP -reseptoren TKV. På grunn av de tilsynelatende konfliktene mellom eksperimentelle studier, gjenstår mye arbeid for å tydeliggjøre rollene til de fysiologiske prosessene som styrer morfogentransport. Et eksempel på at morfogensignalering blir påvirket av ekso- og endocytose, sees i utvikling av Drosophila Air Sac Primordia (ASP), som avhenger av DPP-signalering. Cytoneme-basert signalering bruker mange av de samme komponentene som finnes i nevrale synapser. I Drosophila ASP ("Mottakende celler"), spesialiserte filopodia-lignende cytonemer endocytose DPP fra de tilstøtende Drosophila-vingeskivcellene ("Sendende celler"; figur 3A).
BILDE 6. Vektor transport

Morfogentransportmekanismer.
A, utvidet filopodia kalt Cytonemes er til stede i Drosophila Air Sac Primordium (ASP). Cytonemes som projiserer fra ASP tar opp DPP fra den tilstøtende vinge -forestillingen. Korrelerte transienter av Ca2+ konsentrasjoner blir observert i cytonemer. Cytoneme-mediert transport krever synaptotagmin-4 (SYT4), som hjelper til med vesikkelfusjon og reseptorinternalisering, og glutamatreseptor glurii i ASP. Vingeskivceller som utskiller DPP krever synaptobrevin, synaptotagmin-1, glutamattransportøren og spenningsgatede kalsiumkanaler. Blå sirkler representerer morfogener, røde sirkler representerer PMAD -molekyler i celler, og oransje sirkler representerer DPERK -molekyler i celler. B, i planaria, transporteres morfogener på mikrotubule -matriser langs aksonene fra nervesystemet til sårkanten under regenerering. Vektortransport er identifisert som et grunnleggende krav for skalafri selvsamling av morfogener under planaria homeostase og regenerering. Figuren er tilpasset med tillatelse fra referanse
DPP-signalering ble kompromittert når synaptotagmin-1, synaptobrevin, glutamatreseptoren eller spenningsgaterte Ca2+ -kanaler ble hemmet i de utskillende skivecellene, noe som resulterte i en reduksjon i størrelsen på ASP. Dette resultatet paralleller neurotransmisjon da Synaptobrevin er et iboende membranprotein som regulerer frigjøring av nevrotransmitter gjennom Ca2 +-avhengig eksocytose. Mottakscellene til ASP krever synaptotagmin-4 og glutamatreseptoren glurii. Dette er bemerkelsesverdig gitt at Ca2+ er en avgjørende regulator av endocytose (3) og nevrotransmitterregulering. Huang et al demonstrerte videre at Ca2+ transienter observert i Cytonemes korrelerer med DPP -opptak. Dette antyder at signalopptak og transport i Cytonemes avhenger av lokale Ca2+ -konsentrasjoner. Denne studien understreker dermed rollen som fysiologiske signaler, som Ca2+, i morfogenformidlet transport og internalisering, som ennå ikke er blitt utforsket fullt ut. Ytterligere bevis for andre messenger -signalering og morfogentransport ligger i samspillet mellom Ca2+ og endocytiske og eksocytiske regulatorer. Knockdown av Ca2+ signalavhengige eksocytiske komponenter i den protorakiske kjertelen i Drosophila-hjernen, resulterte i akkumulering av uutgitt steroidhormon ecdyson. Hvorvidt det samme eksocytiske maskineriet kontrollerer sekresjon av viktige morfogener er foreløpig ukjent. Omvendt genetisk RNAi -screening kan brukes for å kartlegge komponentene som regulerer morfogensekresjon og transport gjennom eksocytose. I tillegg vil syntese av nye alleler med mutasjoner i Ca2+ bindende domener av viktige morfogen-regulerende proteiner ytterligere muliggjøre bekreftelse av fysiologiske signaler i å regulere morfogensekresjon.
En nøkkelaktør i den endocytiske prosessen er den regulatoriske guanosin trifosfatase (GTP) protein Rab5. Rab5 -proteiner hjelper til med dannelse av transportvesikler og regulerer nedbrytning av molekylær last og resirkulering. Rab5 er nødvendig for endosomintegritet i den presynaptiske terminalen i Drosophila neuromuskulære synapser. Nedsatt Rab5-funksjon påvirker både ekso- og endocytosehastigheter og reduserer evnen til frigjøring av nevrotransmitter, mens overuttrykk av Rab5 øker frigjøringseffekten til nevrotransmitter. Dette er spesielt interessant fordi Ca2+ er en viktig regulator for frigjøring av nevrotransmitter. Nyere arbeid har vist at Ca2+ -kanaler regulerer bulk-endocytose, en form for endocytose av synaptiske vesikler ved nerveterminaler, i tillegg til kobling av ekso- og endocytose. Videre krever Rab5-avhengig endocytose Ca2+ signalering for å øke hastigheten på membrankapasitans, som bestemmer hvor raskt membran-potensialet kan svare på en endring i strøm, og er lineært relatert til endringer i membranoverflatearealet. Omvekten ble også vist der lave Ca2+ -konsentrasjoner reduserte membrankapasitansen. Dermed reflekterer en økning i overflatearealet økt eksocytose, og redusert overflateareal reflekterer en økning i endocytose. I tillegg viste oppdagelse av en tilbakemeldingssløyfe BMP og Scrib, et stillasprotein assosiert med cellulær spredning, fremmer Rab5 endosomavhengig BMP/DPP-signalering under morfogenese i Drosophila. Gitt at Ca2+ påvirker Rab5-relatert endocytose og Rab5 påvirker DPP-signalering, kan det utledes at Ca2+ -konsentrasjoner kan tjene som en potensiell modulator for morfogentransport gjennom koblet ekso- og endocytose. Imidlertid gjenstår denne slutningen å testes direkte. En annen mulig mekanisme som fysiologiske signaler som Ca2+ påvirker transport av morfogener er gjennom Ca2+ bindende domener til transport-proteiner. Bevis for dette ligger i nærvær av Ca2+ bindende EF-domener i proteiner involvert i dannelsen av BMP- og DPP-gradienter i Drosophila-vingeskiven. For eksempel samhandler Drosophila Pentagone (Pent) direkte med Dally for å gi lang rekkevidde-distribusjon av DPP -liganden. Struktur
Etter dette benyttet Thomas et al SMOC-slettingsmutantkonstruksjonene XSMOC-1LambdaEC (mangler det ekstracellulære Ca2+ -bindingsdomenet) og XSMOC-1EC (som bare inneholder det ekstracellulære Ca2+ bindingsdomenet) for å demonstrere at normal XSMOC-1 og XSMOC1LambdaEC, men ikke XSMOC-MOS-1 1EC konverterer skjebnen til naive Xenopus ektodermeksplanter til fremre nevralt vev. Således krever ikke embryonale celle skjebnebeslutninger om å bli nevrale vev via SMOC, og BMP -hemming krever ikke det ekstracellulære Ca2+ bindingsdomenet. Potensieringen av BMP-signalering av det ekstracellulære Ca2+ -bindingsdomenet ble imidlertid vist i videre studier ved å observere affinitetene til XSMOC-1EC og BMP2 for hverandre og andre heparansulfatproteoglykaner (HSPG). Eksperimenter som benyttet in vitro diffusjonsanalyser på agarosegeler innebygd med eller uten HSPG -er demonstrerte at bindingen av BMPs til HSPG -er begrenser deres virkningsområde og at BMP -diffusjon kan forbedres ved binding av SMOC til HSPG for å utvide virkningsområdet. SMOC -ekstracellulære Ca2+ bindingsdomenet utvider rekkevidden for BMP -signalering gjennom konkurrerende binding til HSPG -er. Disse studiene viser viktigheten av SMOC -ekstracellulært Ca2+ bindingsdomenet i sammenheng med utvikling. Ytterligere undersøkelser er nødvendige i rollen som fysiologisk signalering innenfor konteksten av det ekstracellulære miljøet.
Koblingen mellom neuronale og ikke-nevrale vev for å definere posisjonsinformasjon strekker seg utover likheter i signalmekanismer. En fersk studie ga ytterligere innsikt i nervesystemets rolle i morfogenbasert mønsterdannelse og regenerering i planaria. Et bemerkelsesverdig trekk ved regenerering i Planaria er reformasjonen av viktige morfogengradienter som HH og Notum Regulating Factor (NRF) etter fragmentering eller såring av organismen. Pietak et al utviklet en kvantitativ modell for å regenerere planaria for å belyse mekanismen for morfogengradientaktivitet som sikrer robust kroppsplanregulering. Modellen spår brøkdelen av heteromorfoser i regenererende planariafragmenter. Gjennom en iterativ kombinasjon av beregningssimuleringer og eksperimenter fant de at vektortransport av morfogener var nødvendig for å forklare regenerering av mønsterdannelse. Morfogenvektorbasert transport er definert som retningsstransport av morfogener av et vektorfelt. Vektortransportfeltet falt sammen med nervepolariteten gjennom å regenerere planarisk vev. I sin Markov -kjedemodell er transporten av morfogener, som HH og NRF, formidlet av kinesin og dyneinmotoriske proteiner langs mikrotubulens nettverk av aksoner (figur B ovenfor)
De demonstrerte videre at hodehaleaksen styres av nevronens nettopolaritet. I kontrast kunne en rent diffusjonsbasert modell for mønstring ikke forklare skaleringen av jevn konsentrasjoner av morfogener av fragmenter i forskjellige størrelser. Videre ville diffusjon være for treg til å regenerere mønsteret (krever mer enn en uke på en 1 cm skala organisme), mens Planaria kan reformere innen 72 timer eller mindre. Dermed spiller nervesystemet en lærerikt rolle i å regulere langdistanse aksial mønstringsreparasjon under regenerering.
Bioelektriske signaler og hvilemembranpotensialer over vev er avgjørende for riktig mønster i flere organismer. Endringer i membranpotensial i regenerering av planaria, indusert av ionoforbehandling, permanent påvirket genuttrykk, mønstring og polaritet. Etter ionophore utvasking fra det behandlede vevet, vedvarte de induserte endringene i hvilemembranpotensialet. Denne mekanismen parallellerer synaptisk plastisitet i hjernen der handlingspotensialer, modulert av spenning-gatede ionekanaler, forplanter signaler. Arbeid utført i Xenopus og Chick gir innsikt i denne forekomsten der serotonin (5-HT), en endogen nevrotransmitter, etablerer mønster fra venstre til høyre i embryoer gjennom regulering av ionestrømmer. En oppfølgingsstudie viste at ekstracellulær 5-HT-tilgjengelighet driver innervasjon gjennom vev via gap-junctional kommunikasjonsmodulasjon. Ved å kombinere funnene deres, foreslår Levin og kolleger at ekstracellulær 5-HT, et positivt ladet molekyl, navigerer gjennom gapskryss for å akkumulere i hyperpolariserte celler, og etterligne gjenopptaksfunksjonen til 5-HT-transportører (SERTS). Disse 5-HT-sekvestering, hyperpolariserte celler kan depolarisere som respons på tap av kloridioner via glycin-gated kloridkanalaktivering. Uten normal hyperpolarisering eksporteres 5-HT til det ekstracellulære rommet av SERTS, som kan indusere vekst og hyperinnervasjon av vev. Betydningen av ionekanaler for å lette kommunikasjon i bakterielle biofilmsamfunn viser generaliteten av fysiologisk signalering for å formidle celle-til-cellekommunikasjonsmekanismer. For eksempel bruker bakterielle biofilmsamfunn synkroniserte svingninger av kortdistansekobling blant noen få celler og samfunnsdekkende signalering, for å minimere konkurransen om ressurser. I sum er det nødvendig med ytterligere kvantitative undersøkelser for å utforske rollen som nevrotransmisjon, Gap Junction -kommunikasjon og nervesystemet i morfogenbaserte transportsystemer. Etablering av morfogengradienter gjennom nevral signalering må fremdeles integreres med kjente fysiologiske regulatorer for mønstring. Dette vil være kritisk for å vurdere hvordan mønstre dannes på tvers av store romlige domener der diffusjonsbaserte mekanismer blir ineffektive.
Fysiologiske roller av Ca2+ i formidling av morfogenrespons.
A, en gradient av Morfogen Sonic Hedgehog (SHH) leder mønstringen av neuronal differensiering i den utviklende ryggmargen. SHH øker Ca2+ piggaktivitet i utvikling av ryggmargsneuroner. En gjeldende modell antyder at Ca2+ pigger konverterer støyende HH -signalering til binære utganger for å spesifisere cellens skjebner. Dette ble demonstrert ved en positiv sammenheng mellom aktivering av pinnsvin -signalering gjennom aktivering av SHH -transduseren glattet (SMO) og stimulerende Ca2+ piggaktivitet. Tap av Ca2+ piggene resulterte i reduserte GABAergiske neuronale celle -skjebner. Dette antyder Ca2+ piggfrekvens koder for SHH -konsentrasjon og er nødvendig for riktig neuronal differensiering.
B, vevsdekkende, langdistanse Ca2+ svingninger er blitt observert i mesenkymale celler. Synergistiske handlinger av SHH og Wnt -signalering tillot synkroniserte Ca2+ svingninger for å koordinere cellebevegelser under kyllingfjærforlengelse.
Bilde 7.Celle-differensiering og fjær-utvekst
 I tillegg til å formidle cellulær differensiering og spredning, påvirker Ca2+ -signaler også cellulær migrasjon som respons på morfogengradienter. Koordinert cellemigrasjon under forlengelse av kyllingfjær er ledsaget av dynamiske endringer av bioelektriske strømmer og Ca2+ signalering. Spesifikt inneholdt SHH-responsive celler synkroniserte Ca2+ svingninger der SHH spiller en nøkkelrolle i å formidle interaksjoner mellom epitel og mesenchym under fjærmorfogenese. Spenningsgatede Ca2+ -kanaler og Connexin43-baserte gapskryss regulerer Ca2+ -dynamikk under fjærforlengelse. SHH-signalering og beta-catenin-signalering aktiverer Connexin-43-uttrykk transkripsjonelt for å etablere GAP Junctional Network (figur B). Etableringen av gap -kryssnettverk i voksende vev endrer således det romlige området for Ca2+ signalering
I tillegg til å formidle cellulær differensiering og spredning, påvirker Ca2+ -signaler også cellulær migrasjon som respons på morfogengradienter. Koordinert cellemigrasjon under forlengelse av kyllingfjær er ledsaget av dynamiske endringer av bioelektriske strømmer og Ca2+ signalering. Spesifikt inneholdt SHH-responsive celler synkroniserte Ca2+ svingninger der SHH spiller en nøkkelrolle i å formidle interaksjoner mellom epitel og mesenchym under fjærmorfogenese. Spenningsgatede Ca2+ -kanaler og Connexin43-baserte gapskryss regulerer Ca2+ -dynamikk under fjærforlengelse. SHH-signalering og beta-catenin-signalering aktiverer Connexin-43-uttrykk transkripsjonelt for å etablere GAP Junctional Network (figur B). Etableringen av gap -kryssnettverk i voksende vev endrer således det romlige området for Ca2+ signalering
En gjennomgang av rollen som gapskryss i regulering av mønsterdannelsesdetaljer som venstre-høyre asymmetri i C. elegans nevroner involverer Ca2+ signalering og kommunikasjon gjennom gap-veikryss.94,95 Disse studiene viser videre rollen som andre messenger-systemer i formidling Morfogeninduserte responser av celler og fysiologisk signalering i morfogenbasert mønsterdannelse. Kvantitative eksperimenter som undersøker samspillet mellom andre budbringere og gapskryss under mønsterdannelse er sterkt nødvendig.
Fysiologiske signaler modulerer morfogenbaserte posisjonsinformasjonssystemer
Ca2+ signalering spiller også en nøkkelrolle i å formidle cellulære responser på morfogener. Flere studier i forskjellige modellsystemer har vist samspillet mellom HH -signalering og Ca2+ -dynamikk. For eksempel viste Shaw et al at intracellulær Ca2+ -mobilisering regulerer nivået av sonisk pinnsvin (SHH) -avhengige ekspresjonsdomener til NKX2.2B, ISL1, NKX6.1 og PAX3-gener i det utviklende nervesystemet med zebrafishembryoer under 18- Somitt scene. Videre viste de at redusert ekspresjon av ryanodinreseptor (RYR), en intracellulær Ca2+-frigjøringskanal, forskjøvet tildelingen av SHH-avhengige cellefatter i den somittiske muskelen og nevrale rør på en måte som lignet effekten av redusert SHH-signalering. Disse funnene ble oppdaget ved å bruke tap av funksjonsmutasjoner, antisense morpholino knockdowns og farmakologiske behandlinger for å forstyrre RYR-aktiviteten.
I en annen studie demonstrerte Belgacem og kolleger at SHH akutt øker Ca2+ gjennom aktiveringen av transduseren glattet (SMO), som deretter rekrutterer heterotrimeriske GTP-bindende proteinavhengige veier. De demonstrerte at SHH øker Ca2+ piggaktiviteten ved å utvikle ryggmargsneuroner og foreslå at Ca2+ piggfrekvens koder for SHH -konsentrasjon og er nødvendig for riktig neuronal differensiering (figur A).
Morfogentransportmekanismer.
A, utvidet filopodia kalt Cytonemes er til stede i Drosophila Air Sac Primordium (ASP). Cytonemes som projiserer fra ASP tar opp DPP fra den tilstøtende vinge -forestillingen. Korrelerte transienter av Ca2+ konsentrasjoner blir observert i cytonemer. Cytoneme-mediert transport krever synaptotagmin-4 (SYT4), som hjelper til med vesikkelfusjon og reseptorinternalisering, og glutamatreseptor glurii i ASP. Vingeskivceller som utskiller DPP krever synaptobrevin, synaptotagmin-1, glutamattransportøren og spenningsgatede kalsiumkanaler. Blå sirkler representerer morfogener, røde sirkler representerer PMAD -molekyler i celler, og oransje sirkler representerer DPERK -molekyler i celler. B, i planaria, transporteres morfogener på mikrotubule -matriser langs aksonene fra nervesystemet til sårkanten under regenerering. Vektortransport er identifisert som et grunnleggende krav for skalafri selvsamling av morfogener under planaria homeostase og regenerering. Figuren er tilpasset med tillatelse fra referanse
DPP-signalering ble kompromittert når synaptotagmin-1, synaptobrevin, glutamatreseptoren eller spenningsgaterte Ca2+ -kanaler ble hemmet i de utskillende skivecellene, noe som resulterte i en reduksjon i størrelsen på ASP. Dette resultatet paralleller neurotransmisjon da Synaptobrevin er et iboende membranprotein som regulerer frigjøring av nevrotransmitter gjennom Ca2 +-avhengig eksocytose. Mottakscellene til ASP krever synaptotagmin-4 og glutamatreseptoren glurii. Dette er bemerkelsesverdig gitt at Ca2+ er en avgjørende regulator av endocytose 3 og nevrotransmitterregulering. Huang et al demonstrerte videre at Ca2+ transienter observert i Cytonemes korrelerer med DPP -opptak. Dette antyder at signalopptak og transport i Cytonemes avhenger av lokale Ca2+ -konsentrasjoner. Denne studien understreker dermed rollen som fysiologiske signaler, som Ca2+, i morfogenformidlet transport og internalisering som ennå ikke er blitt utforsket fullt ut. Ytterligere bevis for andre messenger -signalering og morfogentransport ligger i samspillet mellom Ca2+ og endocytiske og eksocytiske regulatorer. Knockdown av Ca2+ signalavhengige eksocytiske komponenter i den protorakiske kjertelen i Drosophila-hjernen, resulterte i akkumulering av uutgitt steroidhormon ecdyson. Hvorvidt det samme eksocytiske maskineriet kontrollerer sekresjon av viktige morfogener er foreløpig ukjent. Omvendt genetisk RNAi -screening kan brukes for å kartlegge komponentene som regulerer morfogensekresjon og transport gjennom eksocytose. I tillegg vil syntese av nye alleler med mutasjoner i Ca2+ bindende domener av viktige morfogenregulerende proteiner ytterligere muliggjøre bekreftelse av fysiologiske signaler i å regulere morfogensekresjon.
En nøkkelaktør i den endocytiske prosessen er den regulatoriske guanosin trifosfatase (GTP) protein Rab5. Rab5 -proteiner hjelper til med dannelse av transportvesikler og regulerer nedbrytning av molekylær last og resirkulering. Rab5 er nødvendig for endosomintegritet i den presynaptiske terminalen i Drosophila neuromuskulære synapser. Nedsatt Rab5-funksjon påvirker både ekso- og endocytosehastigheter og reduserer evnen til frigjøring av nevrotransmitter, mens overuttrykk av Rab5 øker frigjøringseffekten til nevrotransmitter. Dette er spesielt interessant fordi Ca2+ er en viktig regulator for frigjøring av nevrotransmitter. Nyere arbeid har vist at Ca2+ -kanaler regulerer bulk endocytose, en form for endocytose av synaptiske vesikler ved nerveterminaler, i tillegg til kobling av ekso- og endocytose. Videre krever Rab5-avhengig endocytose Ca2+ signalering for å øke hastigheten på membrankapasitans, som bestemmer hvor raskt membranpotensialet kan svare på en endring i strøm og er lineært relatert til endringer i membranoverflatearealet. Omvekten ble også vist der lave Ca2+ -konsentrasjoner reduserte membrankapasitansen. Dermed reflekterer en økning i overflatearealet økt eksocytose og redusert overflateareal reflekterer en økning i endocytose. I tillegg viste oppdagelse av en tilbakemeldingssløyfe BMP og Scrib, et stillasprotein assosiert med cellulær spredning, fremmer Rab5 endosomavhengig BMP/DPP-signalering under morfogenese i Drosophila. Gitt at Ca2+ påvirker Rab5-relatert endocytose og Rab5 påvirker DPP-signalering, kan det utledes at Ca2+ -konsentrasjoner kan tjene som en potensiell modulator for morfogentransport gjennom koblet ekso- og endocytose. Imidlertid gjenstår denne slutningen å testes direkte. En annen mulig mekanisme som fysiologiske signaler som Ca2+ påvirker transport av morfogener er gjennom Ca2+ bindende domener til transportproteiner. Bevis for dette ligger i nærvær av Ca2+ bindende EF-domener i proteiner involvert i dannelsen av BMP- og DPP-gradienter i Drosophila-vingeskiven. For eksempel samhandler Drosophila Pentagone (Pent) direkte med Dally for å gi lang rekkevidde distribusjon av DPP -liganden.
Strukturelt har Pent en lignende domenesammensetning, som den for humant SMOC -protein, og både Pent og SMOC -proteiner inneholder Ca2+ bindende EF -domener. Xenopus SMOC-1 (XSMOC-1) protein fungerer som en BMP-antagonist i Xenopus-embryoer selv i nærvær av konstitutivt aktiv BMP-reseptor.84 Ytterligere analyse antyder at SMOC-1 antagoniserer BMP-signalering nedstrøms for reseptorbinding gjennom aktivering av MAPK-signalering. Senere studier demonstrerte evnen til Drosophila-spesifikk Pent til å hemme på lignende måte BMP-signalering i Xenopus nedstrøms for BMP-reseptoren, etter injeksjon av syntetisk Pent mRNA i Xenopus embryoer.
Etter dette benyttet Thomas et al SMOC-slettings-mutant-konstruksjonene XSMOC-1LambdaEC (mangler det ekstracellulære Ca2+ -bindingsdomenet) og XSMOC-1EC (som bare inneholder det ekstracellulære Ca2+ bindingsdomenet) for å demonstrere at normal XSMOC-1 og XSMOC1LambdaEC, men ikke XSMOC-MOS-1 1EC konverterer skjebnen til naive Xenopus ektodermeksplanter til fremre nevralt vev. Således krever ikke embryonale celle skjebnebeslutninger om å bli nevrale vev via SMOC, og BMP -hemming krever ikke det ekstracellulære Ca2+ bindingsdomenet. Potensieringen av BMP-signalering av det ekstracellulære Ca2+ -bindingsdomenet ble imidlertid vist i videre studier ved å observere affinitetene til XSMOC-1EC og BMP2 for hverandre og andre heparansulfat-proteoglykaner (HSPG). Eksperimenter som benyttet in vitro, diffusjonsanalyser på agarosegeler innebygd med eller uten HSPG -er demonstrerte at bindingen av BMPs til HSPG -er begrenser deres virkningsområde og at BMP -diffusjon kan forbedres ved binding av SMOC til HSPG for å utvide virkningsområdet. SMOC -ekstracellulære Ca2+ bindings-domenet utvider rekkevidden for BMP -signalering gjennom konkurrerende binding til HSPG -er. Disse studiene viser viktigheten av SMOC -ekstracellulært Ca2+ bindingsdomenet i sammenheng med utvikling. Ytterligere undersøkelser er nødvendige i rollen som fysiologisk signalering, innenfor konteksten av det ekstracellulære miljøet.
Koblingen mellom nevronale og ikke-nevrale vev for å definere posisjonsinformasjon strekker seg utover likheter i signalmekanismer. En fersk studie ga ytterligere innsikt i nervesystemets rolle i morfogenbasert mønsterdannelse og regenerering i planaria. Et bemerkelsesverdig trekk ved regenerering i Planaria er reformasjonen av viktige morfogen-gradienter som HH og Notum Regulating Factor (NRF) etter fragmentering eller såring av organismen. Pietak et al utviklet en kvantitativ modell for å regenerere planaria for å belyse mekanismen for morfogengradientaktivitet, som sikrer robust kroppsplanregulering. Modellen spår brøkdelen av heteromorfoser i regenererende planariafragmenter. Gjennom en iterativ kombinasjon av beregningssimuleringer og eksperimenter fant de at vektortransport av morfogener var nødvendig for å forklare regenerering av mønsterdannelse. Morfogenvektorbasert transport er definert som retningsstransport av morfogener av et vektorfelt. Vektortransportfeltet falt sammen med nervepolariteten gjennom å regenerere planarisk vev. I sin Markov -kjedemodell er transporten av morfogener, som HH og NRF, formidlet av kinesin og dyneinmotoriske proteiner, langs mikrotubulens nettverk av aksoner (figur B ovenfor)
De demonstrerte videre at hode-hale aksen styres av nevronens nettopolaritet. I kontrast kunne en rent diffusjonsbasert modell for mønstring ikke forklare skaleringen av jevn konsentrasjoner av morfogener av fragmenter i forskjellige størrelser. Videre ville diffusjon være for treg til å regenerere mønsteret (krever mer enn en uke på en 1 cm skala organisme), mens Planaria kan reformere innen 72 timer eller mindre. Dermed spiller nervesystemet en lærerikt rolle i å regulere langdistanse aksial mønstringsreparasjon under regenerering.
Bioelektriske signaler og hvilemembran-potensialer over vev er avgjørende for riktig mønster i flere organismer. Endringer i membranpotensial i regenerering av planaria, indusert av ionoforbehandling, permanent påvirket genuttrykk, mønstring og polaritet. Etter ionophore utvasking fra det behandlede vevet, vedvarte de induserte endringene i hvilemembranpotensialet. Denne mekanismen parallellerer synaptisk plastisitet i hjernen der handlingspotensialer, modulert av spenning-gate ionekanaler, forplanter signaler. Arbeid utført i Xenopus og Chick gir innsikt i denne forekomsten der serotonin (5-HT), en endogen nevrotransmitter, etablerer mønster fra venstre til høyre i embryoer gjennom regulering av ionestrømmer. En oppfølgingsstudie viste at ekstracellulær 5-HT-tilgjengelighet driver innervasjon gjennom vev via gap-junctional kommunikasjonsmodulasjon. Ved å kombinere funnene deres, foreslår Levin og kolleger at ekstracellulær 5-HT, et positivt ladet molekyl, navigerer gjennom mellomrom-kryss for å akkumulere i hyperpolariserte celler, og etterligne gjenopptaksfunksjonen til 5-HT-transportører (SERTS). Disse 5-HT-sekvestering, hyperpolariserte celler, kan depolarisere som respons på tap av kloridioner via glycin-gate kloridkanal-aktivering. Uten normal hyperpolarisering eksporteres 5-HT til det ekstracellulære rommet av SERTS, som kan indusere vekst og hyperinnervasjon av vev. Betydningen av ionekanaler for å lette kommunikasjon i bakterielle biofilm-samfunn viser generaliteten av fysiologisk signalering for å formidle celle-til-celle kommunikasjonsmekanismer. For eksempel bruker bakterielle biofilmsamfunn, synkroniserte svingninger av kortdistansekobling blant noen få celler og samfunnsdekkende signalering, for å minimere konkurransen om ressurser. I sum er det nødvendig med ytterligere kvantitative undersøkelser for å utforske rollen som nevrotransmisjon, kryss -kommunikasjon og nervesystemet i morfogenbaserte transportsystemer. Etablering av morfogengradienter gjennom nevral signalering må fremdeles integreres med kjente fysiologiske regulatorer for mønstring. Dette vil være kritisk for å vurdere hvordan mønstre dannes på tvers av store romlige domener, der diffusjonsbaserte mekanismer blir ineffektive.
Fysiologiske signaler modulerer morfogenbaserte posisjonsinformasjonssystemer
Ca2+ signalering spiller også en nøkkelrolle i å formidle cellulære responser på morfogener. Flere studier i forskjellige modellsystemer har vist samspillet mellom HH -signalering og Ca2+ -dynamikk. For eksempel viste Shaw et al at intracellulær Ca2+ -mobilisering regulerer nivået av Sonic HedgeHog (SHH) -avhengige ekspresjonsdomener til NKX2.2B, ISL1, NKX6.1 og PAX3-gener i det utviklende nervesystemet med zebrafisk-embryoer under 18- Somitt scene. Videre viste de at redusert ekspresjon av ryanodinreseptor (RYR), en intracellulær Ca2+-frigjøringskanal, forskjøvet tildelingen av SHH-avhengige cellefatter i den somittiske muskelen og nevrale rør på en måte som lignet effekten av redusert SHH-signalering. Disse funnene ble oppdaget ved å bruke tap av funksjons-mutasjoner, antisense morpholino knockdowns og farmakologiske behandlinger for å forstyrre RYR-aktiviteten.
I en annen studie demonstrerte Belgacem og kolleger at SHH akutt øker Ca2+ gjennom aktiveringen av transduseren glattet (SMO), som deretter rekrutterer heterotrimeriske GTP-bindende proteinavhengige veier. De demonstrerte at SHH øker Ca2+ piggaktiviteten ved å utvikle ryggmargsneuroner og foreslå at Ca2+ piggfrekvens koder for SHH -konsentrasjon og er nødvendig for riktig neuronal differensiering (figur A -under).
Figur A. Nevral tube og kylling embryo
Fysiologiske roller av Ca2+ i formidling av morfogenrespons.
A) en gradient av Morfogen Sonic Hedgehog (SHH) leder mønstringen av neuronal differensiering i den utviklende ryggmargen. SHH øker Ca2+ piggaktivitet i utvikling av ryggmargsneuroner. En gjeldende modell antyder at Ca2+ pigger konverterer støyende HH -signalering til binære utganger for å spesifisere cellens skjebner. Dette ble demonstrert ved en positiv sammenheng mellom aktivering av HedgeHog -signalering gjennom aktivering av SHH -transduseren glattet (SMO) og stimulerende Ca2+ piggaktivitet. Tap av Ca2+ piggene resulterte i reduserte GABAergiske neuronale celle -skjebner. Dette antyder Ca2+ piggfrekvens koder for SHH -konsentrasjon, og er nødvendig for riktig neuronal differensiering.
B) vevsdekkende, langdistanse Ca2+ svingninger er blitt observert i mesenkymale celler. Synergistiske handlinger av SHH og Wnt -signalering tillot synkroniserte Ca2+ svingninger for å koordinere cellebevegelser under kyllingfjærforlengelse
I tillegg til å formidle cellulær differensiering og spredning, påvirker Ca2+ -signaler også cellulær migrasjon som respons på morfogengradienter. Koordinert cellemigrasjon under forlengelse av kyllingfjær er ledsaget av dynamiske endringer av bioelektriske strømmer og Ca2+ signalering. Spesifikt inneholdt SHH-responsive celler, synkroniserte Ca2+ svingninger, der SHH spiller en nøkkelrolle i å formidle interaksjoner mellom epitel og mesenchym under fjærmorfogenese. Spenningsgatede Ca2+ -kanaler og Connexin43-baserte kryss, regulerer Ca2+ -dynamikk under fjærforlengelse. SHH-signalering og beta-catenin-signalering aktiverer Connexin-43-uttrykk transkripsjonelt for å etablere GAP Junctional Network (figur B). Etableringen av gap -kryssnettverk i voksende vev endrer således det romlige området for Ca2+ signalering.
En gjennomgang av rollen som gapskryss i regulering av mønsterdannelsesdetaljer som venstre-høyre asymmetri i C. elegans nevroner involverer Ca2+ signalering og kommunikasjon gjennom gap-veikryss.(94,95) Disse studiene viser videre rollen som andre messenger-systemer i formidling Morfogeninduserte responser av celler og fysiologisk signalering i morfogenbasert mønsterdannelse. Kvantitative eksperimenter som undersøker samspillet mellom andre budbringere og gapskryss under mønsterdannelse er sterkt nødvendig.
Tilbakemelding fra fysiologisk signalering sikrer robusthet av morfogenbasert signalering.
Viktige aspekter ved morfogengradienter som en kilde til posisjonsinformasjon er robusthet i nærvær av genetisk eller miljømessig støy og riktig skalering av morfologiske mønstre med hensyn til størrelse. Robusthet er et allestedsnærværende trekk ved biologiske systemer som sikrer at spesifikke funksjoner i systemet opprettholdes, til tross for eksterne og interne forstyrrelser. Robusthet av posisjonsinformasjon kan påvirke utviklingen betydelig. Flere nyere studier har begynt å belyse mekanismene som styrer mønsterets robusthet. I sammenheng med planarisk regenerering ble det for eksempel vist at ren reaksjonsdiffusjons-mekanisme av morfogener ikke klarer å gi skalafrie morfogengradienter. Forfatterne antok at i tillegg til den klassiske mekanismen som involverer diffusjon, er retningsbestemt transport av stoffer gjennom nervesystemet nødvendig for å oppnå skalafri morfogenmønstring og bestemmelse av kroppsaksen.
Hvordan ble den regisseringen oppnådd? Hva om robustheten ikke var der helt fra begynnelsen?
Totalt sett støtter denne studien ideen om skalainvarians i utviklingssystemer der morfogengradienter skaleres riktig, til tross for eksterne og interne forstyrrelser. Til tross for omfattende arbeid i skaleringsmekanismene som regulerer skalering av mønstre på vevs-, organ- og organisme- nivå, gjenstår imidlertid biokjemiske mekanismer som ligger til grunn for mønstring av robusthet, å oppdage regulering av nedstrøms responser ved fysiologiske signaler, også at fysiologiske signaler endrer robustheten i morfogenetisk prosesser. Ca2+ -gradienter genereres langs dorsal-ventral aksen til Drosophila-embryoet. Disse konsentrasjonsgradientene dannes under embryonal stadium 5 med høyere Ca2+ -nivåer på ryggsiden. De viste også at manipulering av Ca2+ -gradienten påvirker spesifikasjonen av amnioserosa, lokalisert dorsalt i Drosophila. Denne studien understreker viktigheten av fysiologiske signalveier som benytter Ca2+ i å bidra til spesifikasjonen av ryggembryonalregionen. Ca2+ gradienter påvirker robusthet av morfogenetiske prosesser. Et enestående spørsmål er om denne mekanismen for spesifikasjon av posisjonsinformasjon, påvirkes av Ca2+ -gradienter i andre modellsystemer.
Bilde 8. Development Gen Regulatory Network (dGRN) hos sjøpølse
 I økende grad akkumuleres bevis for betydelig krysskommunikasjonmellom Ca2+ og ROS mellom endoplasmatisk retikulum, et etablert sted med Ca2+ lagring og mitokondrier, et generasjonssted for ROS.102 spesielt, Ca2+ reduserte ROS fra ROS -generasjonssteder innen mitokondrier under normale forhold og ROS fra ROS -generasjonssteder innen mitokondrier under normale forhold -og forbedret ROS -generering når generasjonssteder ble hemmet av farmakologiske midler. (103) Videre identifiserte kvantitativ in vivo -mikroskopi i Drosophila og sebrafiskembryoer ROS som viktig -avhengig mitokondriell ROS -produksjon korrelert med stedet for actomyosin -kabelmontering. Dette antyder at sårindusert ROS-produksjon fremmer helbredelse i Drosophila og sebrafiskembryoer. Samlet viser disse studiene at fysiologisk signalering, i form av ROS og Ca2+, fungerer for å regulere robusthet av morfogenese. Ytterligere eksperimenter er nødvendige for å belyse hvordan ROS og Ca2+ påvirker robusthet av posisjonsinformasjon, i sammenheng med utvikling, regenerering og sykdom.
I økende grad akkumuleres bevis for betydelig krysskommunikasjonmellom Ca2+ og ROS mellom endoplasmatisk retikulum, et etablert sted med Ca2+ lagring og mitokondrier, et generasjonssted for ROS.102 spesielt, Ca2+ reduserte ROS fra ROS -generasjonssteder innen mitokondrier under normale forhold og ROS fra ROS -generasjonssteder innen mitokondrier under normale forhold -og forbedret ROS -generering når generasjonssteder ble hemmet av farmakologiske midler. (103) Videre identifiserte kvantitativ in vivo -mikroskopi i Drosophila og sebrafiskembryoer ROS som viktig -avhengig mitokondriell ROS -produksjon korrelert med stedet for actomyosin -kabelmontering. Dette antyder at sårindusert ROS-produksjon fremmer helbredelse i Drosophila og sebrafiskembryoer. Samlet viser disse studiene at fysiologisk signalering, i form av ROS og Ca2+, fungerer for å regulere robusthet av morfogenese. Ytterligere eksperimenter er nødvendige for å belyse hvordan ROS og Ca2+ påvirker robusthet av posisjonsinformasjon, i sammenheng med utvikling, regenerering og sykdom.
Analyse av genmodulært nettverk avslører morfogenrettet utviklings robusthet i Drosophila, 30 juni 2020 (4)
Genetisk robusthet er et viktig kjennetegn for å tolerere genetiske eller ikke-genetiske forstyrrelser og sikre fenotypisk stabilitet. Morfogener, en type evolusjonært konserverte (ingen evolusjonsendringer) diffusible molekyler, styrer vevsmønstre på en retningsavhengig eller konsentrasjonsavhengig måte ved å differensielt regulere nedstrøms genuttrykk. Hvorvidt det morfogenrettede genregulerende nettverket har genetisk robusthet, forblir imidlertid unnvikende. I denne studien samlet vi 4217 morfogen-responsive gener langs A-P-aksen til Drosophila-vingeskiver fra RNA-seq-dataene, og grupperte dem i 12 moduler. Ved å bruke matematisk modell på de målte dataene konstruerte vi et genmodulært nettverk (GMN) for å dechiffrere modulens regulatoriske interaksjoner og robusthet i morfogenrettet utvikling. Beregningsanalysene på asymptotisk dynamikk av denne GMN demonstrerte at dette morfogenrettede genmodulære nettverket (GMN) er robust for å tolerere et flertall av genetiske forstyrrelser, som har blitt ytterligere validert ved biologiske eksperimenter. Videre, foruten de genetiske endringene, demonstrerte vi videre at denne morfogenrettede Gen Modulære Nettverk (GMN) godt tåler nongenetiske forstyrrelser (HH-produksjonsendringer) via beregningsanalyser og eksperimentell validering. Derfor indikerer disse funnene tydelig at den morfogenrettede GMN er robust som svar på forstyrrelser, og er viktig for Drosophila for å sikre riktig vevsmønstring i vingeskiven.
Multimodal transkripsjonell kontroll av mønsterdannelse i embryonal utvikling 27. desember 2019 (5)
å forutsi hvordan genuttrykksmønstrene som spesifiserer dyrekroppsplaner oppstår fra interaksjoner mellom transkripsjonsfaktorproteiner og regulatorisk DNA er fortsatt en stor utfordring i fysisk biologi. Mens moduleringen av transkripsjonelle utbrudd har blitt implisert som den primære spaken for å kontrollere genuttrykk, finner vi at dette alene ikke kvantitativt kan rekapitulere mønsterdannelse. I stedet finner vi at mønsteret oppstår gjennom felles virkning av 2 regulatoriske strategier - kontroll av sprengning og kontroll av den totale varigheten av transkripsjonen - som stammer fra forskjellige underliggende molekylære mekanismer. Under embryonal utvikling, tett koreograferte mønstre av genuttrykk - grunne gradienter, skarpe trinn, smale striper - spesifiserte celle-skjebner. Riktig posisjonering, skarphet og amplitude av disse mønstrene av cytoplasmatisk mRNA og protein, sikrer pålitelig bestemmelse av dyrekroppsplaner. å forutsi utviklingsresultater krever en kvantitativ forståelse av flyten av informasjon langs den sentrale dogmen: hvordan input transkripsjonsfaktorer dikterer utgangshastigheten til mRNA -produksjon, hvordan denne frekvensen av mRNA -produksjon dikterer cytoplasmatiske mønstre av mRNA, og hvordan disse mRNA -mønstrene fører til proteintønstre som leverer tilbake til genregulerings-nettverket. Mens sammenhengen mellom transkripsjons-faktorkonsentrasjon og produksjons -mRNA -produksjonshastighet har vært gjenstand for aktiv forskning de siste tre tiårene, har forbindelsen mellom denne utgangshastigheten og de resulterende cytoplasmatiske mønstrene av mRNA forblitt stort sett uutforsket. For eksempel kan en gradert stripe av cytoplasmatisk mRNA i et embryo oppstå som et resultat av radikalt forskjellig transkripsjonell dynamikk på enkeltnukleusnivå
BILDE 9. Nuclei-RNAP A-D

Flere moduser for mønsterdannelse ved enkeltcelle transkripsjonell aktivitet.
(A - D) cytoplasmatiske mRNA -mønstre (A) kan oppstå fra transkripsjonsfaktorer som utøver kontroll over den gjennomsnittlige transkripsjonshastigheten (B), transkripsjonstidsvinduet som dikterer når en kjerne er transkripsjonelt aktiv eller i ro (C), eller fraksjonen av aktive kjerner (D) eller en kombinasjon derav.
Den detaljerte cytoplasmatiske fordelingen av mRNA som lager disse stripene er nøkkelen til overføring av romlig informasjon langs genreguleringsnettet som driver Drosophila -utvikling og forsterker behovet for å utvikle modeller av genregulering som er i stand til å koble kvantitative variasjoner i inngangstranskripsjonsfaktormønster for gradert utgang transkripsjons-hastigheter. Vi fant at alle tre regulatoriske strategier som er skissert i fig. over bidrar kvantitativt til dannelsen av Eve Stripe (2).
Først blir en mindre brøkdel av kjernen aktiv og engasjerer seg i transkripsjon i periferien av stripen enn i sentrum, selv om denne reguleringen av brøkdelen av aktive kjerner, bare gir et mindre bidrag til stripedannelse. For det andre er frekvensen av mRNA -produksjon betydelig forhøyet i midten av stripen. Denne analoge kontrollen av transkripsjonshastigheten er utilstrekkelig for å kvantitativt rekapitulere det cytoplasmatiske mRNA -stripemønsteret. I tillegg til kontrollen av frekvensen av mRNA -produksjon blant kjerner, er det en uttalt regulering av tidsvinduet der Eva -loki var engasjert i transkripsjon over stripen, med de i stripesenteret som uttrykte i omtrent 3 ganger lenger enn de i flankene. Selv om det er verdsatt at gener er transkripsjonelt kompetente for begrensede tidsvinduer under utviklingen, fant vi ut at - når det Primær driver derav. Dermed konkluderer vi med at reguleringen av Eve Stripe 2 er multimodal karakter, med bidrag fra 3 distinkte regulatoriske strategier (fig. B - D ovenfor). Ikke desto mindre kan stripedannelse kvantitativt forklares nesten utelukkende gjennom samspillet mellom 2 distinkte kontrollstrategier: binær kontroll av varigheten av transkripsjonelt engasjement. (Fig. C) og kontroll av middelhastigheten for transkripsjon (fig. B).
For å beskrive Eve Stripe 2 transkripsjonsdynamikk, må vi redegjøre for både de korte, forbigående på perioder diktert av transkripsjonelle utbrudd og et lengre transkripsjonstidsvindu som beskriver perioden som loki engasjerer seg i dette transkripsjonelle utbruddet. En eller flere av disse sprengningsparametrene er underlagt romlig kontrollert regulering.
For å beskrive Eve Stripe 2 transkripsjonsdynamikk, må vi redegjøre for både de korte, forbigående på perioder diktert av transkripsjonelle utbrudd og et lengre transkripsjonstidsvindu som beskriver perioden som loki engasjerer seg i denne transkripsjonelle sprengningen. En eller flere av disse sprengnings-parametrene er underlagt romlig kontrollert regulering.
I Drosophila -utvikling forplantes informasjon som er kodet i en håndfull mødre avsatte protein-gradienter gjennom stadig mer komplekse lag med interagerende gener, og kulminerer med spesifikasjonen av den voksne kroppsplanen. A priori er det flere distinkte regulatoriske strategier på encelle-nivået som er i stand til å generere romlig differensierte mønstre av cytoplasmatisk mRNA, hver med distinkte implikasjoner for arten av de underliggende molekylære prosessene i spill. Den gjennomsnittlige transkripsjonshastigheten er hovedsakelig modulert over embryoet ved å stille inn frekvensen av transkripsjonell sprengning. Det har holdt seg uklart om denne moduleringen av transkripsjonshastigheten (og derved mRNA -produksjon) er den dominerende modaliteten som inputkonsentrasjoner av transkripsjonsfaktorer driver dannelsen av mønstre av genuttrykk, eller om det i stedet ganske enkelt er den mest synlige mekanismen blant flere distinkte kontrollstrategier.
Selv om det blir verdsatt at gener kommer til uttrykk for diskrete tidsvinduer i løpet av utviklingen, er i tilfelle av denne Eve Stripe 2 -reporteren - denne binære transkripsjonelt engasjerte/formal logikken aktivt regulert av transkripsjons-faktorer for å drive mønsterdannelse. Viktig er den tidsmessige komponenten i transkripsjonsregulering i spesifisering av utviklingsresultater. Den begrensede avlesningstiden som er pålagt av korte kjerne-sykluser i tidlige Drosophila -utvikling, plasserer strenge begrensninger for den typen reguleringsarkitektur som kan være ansvarlig for å kjøre observerte mønstre av hunchback -genuttrykk. Pionerfaktoren Zelda spiller en nøkkelrolle i å regulere både tidspunktet og sannsynligheten for transkripsjonell aktivering etter mitose. Arbeidet vårt kompletterer disse tidligere observasjonene ved å utforske nok et aspekt av samspillet mellom timing og transkripsjonell regulering.
Referanser:
Oversatt ved Google og gjennomsyn av Asbjørn E. Lund